Poisson
Un article de Wikipédia, l'encyclopédie libre.
Dans la classification classique les poissons sont des animaux vertébrés aquatiques à sang froid, pourvus de nageoires et dont le corps est le plus souvent couvert d'écailles. On les trouve abondamment aussi bien en eau douce que dans les mers : on trouve des espèces depuis les sources de montagnes (omble de fontaine, goujon) jusqu'au plus profond des océans (grandgousier, Anoplogaster cornuta). Le milieu marin étant moins accessible aux humains, de nombreuses espèces restent encore à découvrir.
Ils sont d'une importance capitale aux humains en tant que nourriture partout dans le monde, qu'ils soient pêchés dans la nature ou élevés en pisciculture. Ils sont aussi exploités à des fins récréatives, avec la pêche et l'aquariophilie, et sont parfois exposés dans de grands aquariums publics. Ils jouent un rôle significatif dans de nombreuses cultures, que ce soit en tant que déités et symboles religieux, ou sujets de livres et de films.
Définition et classification [modifier]
Le terme « poisson » est plus précisément employé pour désigner les chordés non tétrapodes, c'est-à-dire un animal avec une colonne vertébrale possédant des branchies toute sa vie et dont les membres, s'il en possède, sont des nageoires.[1] Il faut noter que les poissons ne forment pas un groupe phylogénétiquement homogène, à l'inverse des oiseaux et des mammifères (voir plus bas).
Un poisson typique est « à sang froid » ; il possède un corps allongé lui permettant de nager rapidement ; il extrait l'oxygène de l'eau en utilisant ses branchies ou un organe respiratoire annexe lui permettant de respirer l'oxygène de l'atmosphère ; il possède deux paires de nageoires, habituellement une ou deux (plus rarement trois) nageoires dorsales, une nageoire anale et une nageoire de queue ; il possède une mâchoire ; il possède une peau généralement recouverte d'écailles ; il pond des œufs, la fécondation peut être interne ou externe.

Il existe de nombreuses formes et tailles de poissons : ce
dragon des mers, proche de l'
hippocampe, se confond avec les algues grâces à ses nageoires.
Chacune de ces caractéristiques comporte toutefois des exceptions. Les thons, les espadons et certaines espèces de requins sont entre sang chaud et sang froid, et peuvent élever leur température corporelle au-dessus de l'eau ambiante.[2] La forme du corps et les performances natatoires varient considérablement, des nageurs très rapides capables de parcourir dix à vingt longueurs de leur corps par seconde (thons, saumons) aux poissons très lents mais mieux manœuvrants comme les anguilles ou les raies) qui ne dépassent pas 0,5 longueur par seconde.[3] Plusieurs groupes de poissons d'eau douce extraient l'oxygène de l'air comme de l'eau en utilisant des organes variés. Les dipneustes possèdent deux poumons similaires à ceux des tétrapodes ; les gouramis ont un « organe labyrinthe » qui fonctionne de la même manière ; les Corydoras extraient l'oxygène par l'estomac ou l'intestin.[4] La forme du corps et la position des nageoires varie énormément, comme en témoigne la différence entre les hippocampes, les lophiiformes, les poissons globes ou les saccopharyngiformes. De même, la surface de la peau peut être nue (murènes) ou couverte d'écailles de différents types : placoïdes (requins et raies), cosmoïdes (cœlacanthes), ganoïdes, cycloïdes et cténoïdes.[5] Certains poissons passent même davantage de temps hors de l'eau que dedans, comme les périophthalmes qui se nourrissent et interagissent entre eux sur des terrains boueux et ne retournent dans l'eau que pour se cacher dans leur terrier.[6]
La taille d'un poisson varie du requin baleine de 16 m au Schindleria brevipinguis d'à peine 8 mm.
Il faut noter que plusieurs types d'animaux aquatiques sont communément appelés « poissons » mais n'en sont pas vis--à-vis de la définition ci-dessus.
Les termes relatifs aux poissons viennent de différentes racines :
Classification [modifier]
À l'opposé des groupes tels que les oiseaux ou les mammifères, les poissons ne forment pas un clade unique : il est paraphylétique, c'est à dire qu'il n'est pas phylogénétiquement homogène : tout clade comprenant tous les poissons comprend aussi les tétrapodes, qui ne sont pas des poissons[7],.[8] Pour cette raison, les groupes comme la « classe Pisces », comme on peut lire dans d'anciennes références, ne sont plus utilisés dans les classifications formelles.
Les poissons sont classés dans les groupes principaux suivants (en grisé, les taxons éteints) :
Certains paléontologistes considèrent que les conodontes sont des chordés et les considèrent comme des poissons primitifs ; voir l'article Vertébré.
Les différents groupes de poissons pris tous ensemble comprennent plus de la moitié des vertébrés connus. Il y a près de 28 000 espèces de poissons existantes (sans compter les espèces disparues), dont près de 27 000 poissons osseux, le reste étant formés d'environ 970 requins, raies et chimères et environ 108 lamproies et myxines.[9] Un tiers de toutes ces espèces est renfermé dans les neuf plus grandes familles, qui sont (des plus grandes aux plus petites) : Cyprinidae, Gobiidae, Cichlidae, Characidae, Loricariidae, Balitoridae, Serranidae, Labridae, et Scorpaenidae. D'un autre côté, environ 64 familles sont monotypiques (ne contiennent qu'une seule espèce). On prédit que le nombre total d'espèces de poissons en existence serait de 32 500.[9]
Cliquez pour dérouler
Place des poissons dans le règne animal
Le monde animal :
- Les types d'organisation présentés ici sont des grades évolutifs ne correspondant généralement pas à des groupes monophylétiques, mais paraphylétiques (ne comportant pas tous les descendants d'un même ancêtre – exemple : les descendants d'ancêtres vermiformes ne sont pas tous aujourd'hui des vers, etc.).
- En jaune : les principales explosions radiatives.
|
| >Unicellulaires procaryotes (cellule sans noyau) |
|
♦ Echinodermes: Oursin, Crinoïdes, Concombres de mer, étoile de mer et ophiure. |
|
♦ Bivalves (coquillages) |
|
|
| >Unicellulaires Eucaryotes (cellule à noyau) |
|
|
|
♦ Gastéropodes (escargots, limaces,...) |
|
|
| >Éponge (organisme multicellulaire) |
|
♦ Mollusques |
|
♦ Céphalopodes (Pieuvres, seiches) |
|
|
| >Polype : hydres, corail et méduse |
|
|
|
|
|
|
| >Vers (mobilité et tube digestif) bilatérien |
|
|
|
♦ Trilobites (de deux à 24 pattes - éteint) |
|
|
| >Poissons Agnathes (sans mâchoire) |
|
♦ Arthropodes primitifs type myriapodes (beaucoup de pattes) |
|
♦ Décapodes: crabes et écrevisses (dix pattes) |
|
|
| >Poissons primitifs (poissons cartilagineux) |
|
|
|
♦ Arachnides : araignées, scorpions et acariens (huit pattes) |
|
♦ Libellules |
| >Poissons typiques (poissons osseux) |
|
♦ Serpents |
|
> hexapodes (à six pattes) : Insectes type Apterygota (primitifs sans ailes). |
|
♦ Blattes, mantes, termites. |
| >Poissons type Sarcopterygii (à nageoires charnues) |
|
♦ Dinosaures (éteint) |
|
|
|
♦ Orthoptère (sauterelle, grillon). |
| >Tétrapodes primitifs (type Amphibiens) |
|
♦ Crocodiles |
|
♦ Marsupiaux |
|
♦ Hémiptères (punaises, cigales,...) |
| >Reptiles primitifs (Amniotes de type Lézard) |
|
♦ Tortues |
|
♦ Insectivores (Taupe, Hérisson,...) |
|
♦ Coléoptère (hanneton, coccinelle,...) |
| |
|
♦ Oiseaux |
|
♦ Chiroptères (Chauves-souris) |
|
♦ Hyménoptère (abeille, guêpe, fourmi) |
| |
|
|
|
♦ Primates |
|
♦ Diptère (mouches) |
| |
|
>Mammifères primitifs type monotrème |
|
♦ Rongeurs et Lagomorphes (lapins) |
|
♦ Lépidoptère (papillon) |
| |
|
|
|
♦ Carnivores |
|
♦ |
| |
|
|
|
♦ Ongulés |
|
♦ |
Certains pensent que les poissons ont évolué à partir d'une créature du type ascidie (dont les larves ont des ressemblances avec les poissons primitifs) ; les premiers ancêtres des poissons auraient alors conservé leur forme larvaire à l'état adulte par néoténie, mais l'inverse est aussi possible. Les fossiles candidats au statut de « premier poisson » connus sont Haikouichthys, Myllokunmingia et Pikaia.
Les tout premiers fossiles de poissons ne sont guère nombreux, ni de bonne qualité : peut-être les poissons primitifs étaient-ils rares ou mal fossilisables ou les conditions taphonomiques mauvaises. Cependant, le poisson devint une des formes de vie dominantes du milieu aquatique et a donné naissance aux branches évolutives menant aux vertébrés terrestres comme les amphibiens, les reptiles et les mammifères.
L'apparition d'une mâchoire articulée semble être la raison majeure de la prolifération ultérieure des poissons, car le nombre d'espèces de poissons agnathes devint très faible. Les premières mâchoires ont été trouvées dans les fossiles de placodermes. On ignore si le fait de posséder une mâchoire articulée procure un avantage, par exemple pour la préhension ou la respiration.
La fonctionnalité essentielle qui a initialement structuré cet ordre a été la capacité de nager dans l'eau (ce que ne savaient faire ni les mollusques primitifs, ni les arthropodes primitifs). Toutefois, cette capacité n'a pas conduit à une explosion radiative : par elle-même, elle ne donne pas une autonomie fonctionnelle suffisante pour que les organismes puissent se spécialiser de manière très libre.
L'histoire de cette lignée paraît laborieuse : la découverte progressive de la tête et de la mâchoire, puis l'exploration des membres jusqu'au stade tétrapode, et enfin, la conquête de l'environnement aérien, aboutissant à l'explosion radiative des sauriens.
Les agnathes regroupent des animaux à corde dorsale et à crâne, mais sans mâchoires. Leur vie en milieu aquatique les a fait longtemps classer parmi les poissons.
Le taxon n'est pas considéré comme monophylétique, et doit être considéré comme obsolète dans une classification phylogénétique. Il comprend :
Les myxines et lamproies partagent des caractères morphologiques ancestraux à tous les crâniates, qui seront perdus chez les gnathostomes. Leur bouche rudimentaire, qui se comporte comme une ventouse, ne possède pas de mâchoires, et ne peut donc pas modifier son ouverture. Leur squelette est cartilagineux et composé d'une capsule crânienne et d'une colonne vertébrale sans côtes.
Si les agnathes actuels sont peu nombreux, de nombreux fossiles d'agnathes sont présent dans les sédiments du paléozoïque. Les agnathes furent les tout premiers crâniates à apparaître.
Les fonctionnalités explorées à ce stade évolutif portent sur le fonctionnement de la tête :
- La céphalisation, commune chez tous les vermiformes, s'accentue et le cerveau s'entoure d'un crâne ;
- des branchies apparaissent, de forme variable suivant les groupes ;
- la bouche se dote de dents.
Poissons cartilagineux (Chondrichtyens) [modifier]

Raie photographiée à l'aquarium « Sea Life Helsinki »
On trouve à ce stade évolutif primitif les « poissons cartilagineux » Chondrichthyens, dont le « squelette » est formé de cartilages : les différentes espèces de requins, de raies et de chimères. Le tissu osseux ne sera inventé que plus tardivement, conduisant aux « poissons osseux ».
La principale invention évolutive de ces poissons est la mâchoire et son perfectionnement. En ce qui concerne l'apparition de la mâchoire des gnathostomes (« poissons » à mâchoires, tétrapodes), les os de la paire de branchies la plus antérieure des agnathes semble être homologues des os des mâchoires des gnathostomes.
Ces poissons bénéficient de deux autres acquis qui leur permettent d'être des nageurs plus performants et réactifs : l'oreille interne qui se perfectionne et acquiert un troisième canal semi-circulaire, et surtout les fibres nerveuses qui se couvrent de myéline, ce qui permet une transmission plus rapide de l'influx nerveux.
Cette dernière invention — peu visible mais déterminante — permettra aux vertébrés d'atteindre des tailles importantes, tout en restant nerveux et réactifs. Les « dents de la mer » sont ainsi les premiers à jouer dans la cour des grands, et les grands prédateurs marins actuels prouvent que cette voie évolutive était efficace.
Poissons osseux (Ostéichtyens au sens classique) [modifier]
Comme leur nom l'indique, l'innovation la plus notable des poissons osseux est l'os. Le tissus osseux périchondral qui renforçait certains cartilages se généralise, et conduit à deux types d'os d'origines différentes :
- L'os enchondral (associé au mésoderme), qui remplace au cours du développement les pièces cartilagineuses du squelette interne ;
- surtout, l'os dermique, qui se forme à partir du derme (tissus de l'ectoderme), apparaît. Il donne les os de la boîte crânienne et les ceintures scapulaires, ainsi que les rayons des nageoires (qui évolueront ultérieurement en membres).
On observe aussi la présence de sacs aériens connectés au tube digestif qui donneront les poumons des vertébrés terrestres et les vessies natatoires des Actinoptérygiens. Ces sacs aériens sont soupconnés chez certains Gnathostomes fossiles. Les tentatives d'émancipation du milieu aquatique seraient alors apparues dans ce clade.
Les principales fonctionnalités évolutives explorées au niveau des poissons osseux sont l'articulation de la mâchoire, de plus en plus structurée, et la forme et la mobilité des nageoires.
Poisson à nageoires charnues [modifier]

Dipneuste, dessin d'Heinrich Harder (autour de 1920)
Les poissons à nageoires charnues (sarcoptérygiens) présentent les premiers éléments anatomiques marquant l'évolution vers les tétrapodes :
- le groupe hérite des poissons supérieurs une mâchoire articulée (maxillaire) et les dents se couvrent d'émail ;
- les nageoires paires évoluent vers une forme de membres articulés : l'attache basale se réduit à un seul élément (fémur, omoplate et humérus) et des muscles permettent l'articulation indépendante des rayons (doigts) ;
- le poumon (vessie natatoire / sac à air) devient fonctionnel et muni d'alvéoles.
La dernière évolution du type est la naissance du cou : l'attache du membre antérieur (ceinture scapulaire) se détache de la tête, et les premières vertèbres cervicales se spécialisent pour permettre une articulation du cou.
Les évolutions ultérieures du type sont liées à la sortie du milieu aquatique, et conduisent au groupe des tétrapodes.

Anatomie externe d'un poisson, le
Lampanyctodes hectoris : (1) - opercule (couverture des branchies), (2) - ligne latérale, (3) - nageoire dorsale épineuse, (4) - nageoire dorsale molle (molle ou adipeuse chez les salmonidés), (5) - pédoncule caudal, (6) - nageoire caudale, (7) - nageoire anale, (8) - photophores, (9) - nageoires pelviennes, (10) - nageoires pectorales
Système digestif [modifier]

Bouche et dents d'un
Panaque nigrolineatus en aquarium, d'environ 11 ans.
Chez les poissons, la nourriture est ingérée par la bouche et subit un début de trituration dans l'œsophage. Le broyage des aliments se fait essentiellement dans l'estomac et, chez de nombreuses espèces, dans des diverticules en forme de doigt appelés cæca pyloriques. Ces caeca sécrètent des enzymes digestives et commencent l'absorption des nutriments.
Des organes comme le foie et le pancréas apportent de nouvelles enzymes digestives au fur et à mesure que la nourriture progresse dans le tube digestif. Les intestins complètent le processus de digestion et d'absorption des nutriments. Les poissons mangent des plantes et d'autres organismes.
Système respiratoire [modifier]
Les poissons respirent généralement grâce à des branchies localisées de part et d'autre du pharynx. Les branchies sont constituées de filaments contenant un réseau de capillaires sanguins, dont la grande surface développée facilite l'échange du dioxygène et du dioxyde de carbone.
Les poissons pompent l'eau (contenant du dioxygène) par la bouche, puis la font circuler sur les branchies. Le sang dans les capillaires sanguins s'écoule dans la direction opposée à celle de l'eau, permettant ainsi un échange à contre-courant ; l'eau appauvrie en dioxygène est ensuite expulsée par les ouvertures situées latéralement par rapport au pharynx.
Chez la plupart des poissons cartilagineux (hormis les chimères), les fentes branchiales sont visibles (latéralement chez les requins et les lamproies, ventralement chez les raies). Ces fentes branchiales sont cachées chez les poissons osseux par un opercule (osseux lui-aussi) s'ouvrant postérieurement par une fente appelée ouïe.
De nombreux poissons peuvent respirer de l'air, grâce à différents mécanismes. La peau des anguilles peut être utilisée pour absorber le dioxygène. La cavité buccale des anguilles électriques peut être utilisée pour respirer de l'air. Les poissons-chats des familles Loricariidae, Callichthyidae et Scoloplacidae peuvent absorber l'air à travers leur système digestif.[10] Certaines espèces, tels les dipneustes, possèdent des poumons plus ou moins rudimentaires qui leur permettent de survivre en respirant hors de l'eau dans des mares appauvries en dioxygène ou régulièrement asséchées. Les semionotiformes et le poisson-castor ont une vessie natatoire vascularisée utilisée de la même manière. Les Cobitidae, les trahiras et certains poissons-chats respirent en faisant passer l'air dans l'intestin. Les périophthalmes respirent en absorbant le dioxygène à travers la peau, de façon similaire aux grenouilles. Certains poissons possèdent un « organe de respiration annexe », utilisé pour extraire l'oxygène de l'air ; Les gouramis ont un « organe labyrinthe » au-dessus des branchies pour cela. Quelques autres poissons ont des structures similaires, notamment les familles de poissons-chats Channidae, Luciocephalidae et Clariidae.
La faculté de respirer de l'air est surtout utile aux poissons habitant des eaux peu profondes où la concentration en oxygène peut baisser à certains moments de l'année. Pendant ces périodes, les poissons qui dépendent uniquement de l'oxygène contenu dans l'eau (comme les perches) suffoquent rapidement tandis que les poissons pouvant respirer de l'air survivent mieux, même dans une eau qui ressemble davantage à de la boue humide. Dans des cas extrêmes, certains de ces poissons peuvent survivre plusieurs semaines dans des replis humides, à des endroits où l'eau s'est presque complètement retirée, dans un état d'estivation.
Système circulatoire [modifier]
Les poissons ont un système circulatoire clos comportant une pompe cardiaque. Le circuit sanguin est constitué d'une boucle unique : le sang va du cœur aux branchies, puis au reste du corps, et, finalement, revient au cœur, à l'intérieur duquel ne circule que du sang veineux.
Chez la plupart des poissons, le cœur est divisé en quatre parties :
- le sinus veineux qui collecte le sang veineux et le dirige vers l'atrium ;
- l'atrium, poche musculaire à sens unique qui dirige le sang vers le ventricule ;
- le ventricule cardiaque, poche musculaire qui effectue le travail de pompage cardiaque ;
- le bulbe cardiaque qui, relié à l'artère aorte, permet l'éjection du sang vers les branchies.
Même s'il comporte quatre parties, le cœur d'un poisson comporte deux compartiments[11]
Chez les poissons pulmonés, on remarque un cloisonnement du cœur analogue à celui des tétrapodes pulmonés. La circulation est double, fermée et complète.
Système excréteur [modifier]
Certains déchets métaboliques peuvent diffuser, directement à travers les branchies, du sang vers l'eau environnante. Les autres sont éliminés par les reins, organes excréteurs qui filtrent les déchets pour les éliminer du sang. Comme de nombreux animaux aquatiques, les poissons rejettent leurs déchets azotés sous forme d'ions ammonium (NH4+).
Les poissons d'eau de mer ont tendance à perdre de l'eau à cause des phénomènes d'osmose et les reins concentrent les déchets afin de réabsorber l'eau au maximum. Il se passe l'inverse chez les poissons d'eau douce, qui ont tendance à absorber de l'eau par osmose. Les reins produisent alors de grandes quantités d'urine très diluée.
Certains poissons qui migrent du milieu salé ou milieu dulçaquicole (anadromes, catadromes) ont des reins capables de s'adapter au milieu et de changer leur mode de fonctionnement.
Système nerveux et sensoriel [modifier]
Les poissons ont un système nerveux bien développé qui s'organise autour d'un cerveau divisé en plusieurs parties :
- le bulbe olfactif, dans la partie la plus antérieure, est impliqué dans le sens de l'odorat ;
- le cerveau stricto sensu, contrairement à celui des autres vertébrés, s'occupe plus du traitement des informations concernant le sens de l'odorat que de la réalisation des mouvements volontaires ;
- les lobes optiques traitent les informations provenant des yeux ;
- le cervelet coordonne les mouvements du corps ;
- le bulbe rachidien contrôle le fonctionnement des organes internes.
La plupart des poissons possèdent des organes sensoriels très développés. La majorité des poissons diurnes ont des yeux capables d'une vision des couleurs au moins aussi bonne que celle des humains. De nombreux poissons possèdent aussi des cellules spécialisées appelées chémorécepteurs responsables d'un extraordinaire sens du goût et de l'odorat. Bien qu'ils aient des oreilles internes, de nombreux poissons entendent mal les sons. Cependant, la plupart des poissons possèdent des récepteurs sensitifs formant la ligne latérale, qui leur permet de détecter les courants faibles et les vibrations (par exemple le mouvement de leur proie).[12] Les poissons-chats et les requins possèdent de plus des organes qui détectent des courants électriques de faible intensité. Certaines espèces — telles les mormyridés, notamment Gnathonemus petersii — peuvent être utilisées pour détecter la pollution de l'eau grâce à la variation de leurs émissions électriques. Des chercheurs de l'université d'Édimbourg (Écosse) travaillant sur la truite arc-en-ciel ont découvert en 2003 que les poissons pouvaient ressentir la douleur.[13] Cette étude a été critiquée par James D. Rose la même année,[14] au motif qu'il manque aux poissons un néocortex pour ressentir la douleur. Un article datant de la même année avançait que les poissons ne ressentaient pas la douleur, mais réagissaient simplement aux stimulis.[15]
Système locomoteur [modifier]
La plupart des poissons se déplacent en contractant alternativement les muscles insérés de chaque côté de la colonne vertébrale. Ces contractions font onduler le corps de la tête vers la queue. Lorsque chaque ondulation atteint la nageoire caudale, la force propulsive créée pousse le poisson vers l'avant.
Les nageoires du poisson sont utilisées comme stabilisateurs. La nageoire caudale sert aussi à augmenter la surface de la queue, augmentant ainsi la poussée lors de la nage, et donc la vitesse. Le corps fuselé des poissons permet de diminuer les frictions lorsqu'ils nagent, et donc d'éviter qu'ils soient ralentis par la résistance de l'eau. De plus, leurs écailles sont enrobées d'un mucus qui diminue les frottements.
Puisque les tissus corporels sont plus denses que l'eau, les poissons doivent compenser les effets de la gravité pour éviter de couler. De nombreux téléostéens ont un organe interne appelé vessie natatoire qui ajuste leur flottabilité en réduisant ou augmentant le volume de gaz qu'elle contient, selon le même système que les ballasts des sous-marins.
Remarque : chez certains poissons électriques (raie torpille par exemple), la production d'électricité est réalisée par des cellules musculaires modifiées.
Système reproducteur [modifier]
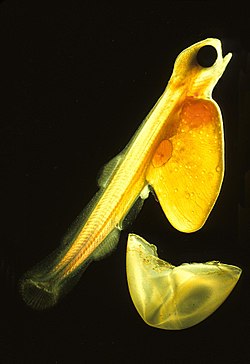
Larve de
saumon ; on distingue les artères autour de la
vésicule vitelline, des gouttes d'huile, le système digestif, la principale artère caudale, la vessie et les arcs des branchies. 24 heures plus tard, il n'aura plus son sac de jaune.
Il existe différents modes de reproduction : l'oviparité, l'ovoviviparité et la viviparité.
Chez les poissons ovipares, après une fécondation généralement externe, la femelle pond ses œufs, et les embryons se développent et éclosent en dehors de son corps. Les embryons sont nourris grâce aux réserves contenues dans l'œuf. Plus de 97 % des espèces connues de poissons sont ovipares,[16] dont par exemple les saumons, les poissons rouges, les thons, les anguilles, etc. Il existe différentes techniques de ponte :
- abandon des œufs au hasard ;
- dépôt des œufs dans un nid construit ou un abri naturel ;
- suspension des œufs à un support ;
- dépôt puis mise à l'abri des œufs dans la bouche d'un des parents ;
- enfouissage des œufs dans les sédiments.
La fécondation est le plus souvent externe, les gamètes se mélangeant à proximité des deux poissons. Chez quelques poissons, la fécondation est interne, le mâle utilisant un organe intromittant pour déposer le sperme dans le cloaque de la femelle. Il s'agit surtout des requins ovipares comme le requin dormeur cornu et des raies ovipares (comme Rajidae). Dans ce cas, le mâle possède deux ptérygopodes, mutations de la nageoire pelvienne.
Les jeunes poissons sortant de l'œuf sont appelés larves. Elles sont généralement peu formées, transportent une vésicule vitelline (ou sac vitellin) contenant leurs réserves nutritives et ont une apparence très différente des adultes. La période larvaire des poissons ovipares est généralement courte (quelques semaines), les larves grandissant rapidement tandis que leur structure et leur apparence change (la métamorphose) pour ressembler aux jeunes de leur espèce. Pendant cette période, les larves se nourrissent d'abord de leurs réserves puis du zooplancton ; ce processus dépend de la concentration en zooplancton et peut causer une forte mortalité parmi les larves.
Chez les poissons ovovivipares, les œufs restent dans le corps de la mère après fécondation interne. Chaque embryon se développe dans son œuf, sans utiliser de réserves nutritives de la mère, puis sort du corps de sa mère lors de l'éclosion. Parmi les poissons ovovivipares, on peut noter les guppys, les squatiniformes et les cœlacanthes.
Chez les poissons vivipares, la fécondation est aussi interne mais chaque embryon reçoit ses nutriments du corps de sa mère, et non des réserves de l'œuf. Les petits sont mis au monde par accouchement. Les poissons vivipares ont une structure similaire au placenta des mammifères, reliant le sang de la mère et celui de l'embryon. Parmi les poissons vivipares, on peut noter les Embiotocidae, les Goodeidae et les requins citrons. Les embryons de certains poissons vivipares ont un comportement appelé oophagie, lorsque l'embryon en développement mange les œufs produits par la mère. Ce comportement a été observé chez certains requins comme le requin mako ou le maraîche mais aussi chez d'autres poissons comme le Nomorhamphus ebrardtii.[17] Le cannibalisme in utero est un comportement encore plus inhabituel, lorsque l'embryon le plus grand mange ses frères plus petits et plus faibles. Ce comportement a été observé chez des requins comme Carcharias taurus mais aussi avec des espèces comme Nomorhamphus ebrardtii.[17]
Système immunitaire [modifier]
Les types d'organes immunitaires varient selon les types de poissons:[18]
- Chez les Agnathes (myxines et lamproies), il n'existe pas de véritables organes immunitaires. En revanche, ces poissons possèdent des tissus lymphoïdes, capables de produire des cellules immunitaires, dans leurs organes. Par exemple, les érythrocytes, macrophages et plasmocytes sont produits dans le rein antérieur (ou pronéphros), et certaines zones de l'intestin (où les granulocytes font leur maturation) chez la myxine ressemble à une moelle osseuse primitive.
- Les poissons cartilagineux (requins et raies) ont un système immunitaire plus évolué que les Agnathes. Ils possèdent trois organes spécialisés qui sont typiques des Chondrichtyens :
- les organes épigonaux, entourant les gonades, qui ressemblent à la moelle osseuse des mammifères ;
- l'organe de Leydig, à l'intérieur des parois de l'œsophage ;
- une valve spirale dans l'intestin.
Tous ces organes contiennent des cellules immunitaires typiques (granulocytes, lymphocytes et plasmocytes). Les Chondrichtyens possèdent aussi un thymus identifiable et une rate bien développée (qui est leur organe immunitaire le plus important) où divers lymphocytes, plasmocytes et macrophages se développent et sont stockés.
- Les Chondrostéens (esturgeons et Polyodontidae) et les polyptères possèdent un site principal de production de granulocytes dans une masse cellulaire associée aux méninges, membranes entourant le système nerveux central), et leur cœur est souvent couvert d'un tissu qui contient des lymphocytes, des fibroblastes et quelques macrophages. Le rein des Chondrostéens est un organe hématopoïétique important où les érythrocytes, granulocytes lymphocytes et macrophages se développent ;
- Les Téléostéens, ou poissons osseux ont, comme les Chondrostéens, leurs principaux tissus immunitaires dans les reins (surtout dans le rein antérieur, où l'on trouve de nombreuses cellules immunitaires de type varié).[19] Les Téléostéens possèdent de plus un thymus, une rate et des zones immunitaires éparses intégrées dans des tissus muqueux (par exemple dans la peau, les branchies, l'intestin et les gonades). On pense que comme chez les mammifères, leurs érythrocytes et granulocytes (entre autres les neutrophiles) résident dans la rate, alors que les lymphocytes sont majoritaires dans le thymus[20],.[21] D'après une découverte récente non encore confirmée, un système lymphatique similaire à celui des mammifères a été décrit chez un Téléostéen, le poisson zèbre (Brachydanio rerio). Ce système serait l'endroit où les lymphocytes T s'accumuleraient en attendant d'être mis en contact avec un antigène.[22]
Température corporelle [modifier]
La plupart des poissons sont des animaux dits à sang froid (poïkilothermes). Plus exactement, leur température interne varie avec celle du milieu (ectothermes).
Des études récentes montrent que certaines espèces prédatrices sont capables de maintenir leur température interne au-dessus de celle de l'eau. Cette aptitude a été démontrée chez certains téléostéens — Scombroïdés (marlin, espadon, voilier), gastéro (Gasterochisma melampus) — et chondrostéens — Lamnidés (requin mako, requin blanc, requins taupes). Leur capacité à réguler leur température varie de l'espadon, qui ne réchauffe que ses yeux et son cerveau, au thon bleu et aux requins taupes capables d'élever leur température corporelle de 20 °C par rapport à celle de l'eau. Des retes mirabiles constituent une sorte d'échangeur thermique et permettent de récupérer les calories du sang chaud et pauvre en oxygène. Le sang qui arrive aux organes est donc relativement chaud tout en diminuant le coût énergétique. Cette méthode accroît la force contractile des muscles, augmente le rendement de la digestion et du système nerveux central, et permet par exemple aux thonidés de chasser en eau profonde durant la nuit et de capturer facilement sardines et maquereaux engourdis par le froid.
Comportement social [modifier]
Un certain nombres d'espèces de poissons, tels les demoiselles (Pomacentridés), les poissons zèbres (Cyprinidés) ou les néons (Characidés), ont un instinct grégaire et préfèrent vivre en bancs. D'autres, tels les requins, sont plutôt solitaires. Certains sont même si agressifs que la rencontre d'un congénère peut entraîner la mort de l'un d'eux après une rude bataille : c'est le cas du combattant du Siam (Anabantidés).
Migration des poissons [modifier]
Menaces et conservation [modifier]
Pour les poissons comestibles comme la morue et le thon, la menace principale est la surpêche[23],.[24] Lorsque la surpêche persiste, elle finit par causer une diminution de la population de poissons (le « stock ») car les individus ne peuvent pas se reproduire assez vite pour compenser la perte dûe à la pêche. Un exemple de surpêche catastrophique très étudié est celui de la sardine du Pacifique (Sadinops sagax caerulues), qui était pêchée près des côtes de Californie. Le maximum avait été atteint en 1937 avec 790 000 tonnes, puis la quantité pêchée a décru pour atteindre à peine 24 000 t en 1968, date à laquelle cette industrie s'arrêta faute d'être rentable. Une telle « extinction commerciale » ne veut pas dire que l'espèce elle-même est éteinte, mais seulement qu'elle n'est plus économiquement viable.[25]
La principale source de tension entre l'industrie de la pêche et la science halieutique est la recherche d'un équilibre entre la conservation des espèces pêchées, et la préservation du revenu des pêcheurs. Dans des zones comme l'Écosse, Terre-Neuve ou l'Alaska, où l'industrie des pêches est le principal employeur, le gouvernement est particulièrement impliqué dans cet équilibre,[26] en maintenant à la fois un stock suffisant et des ressources suffisantes pour les pêcheurs. D'un autre côté, les scientifiques promeuvent une protection toujours accrue pour les stocks, en prévenant que de nombreux stocks pourraient disparaitre dans les cinquante prochaines années.[27]
Destruction des habitats [modifier]
Une des menaces sur les écosystèmes tant marins que d'eau douce et la dégradation de l'habitat ; celle-ci est causée par la pollution de l'eau, la construction de barrages, la diminution du niveau d'eau par la consommation humaine, et l'introduction d'espèces invasives exotiques.[28] Un exemple de poisson en danger à cause d'un habitat modifié est l'esturgeon Scaphirhynchus albus, vivant dans les cours d'eau en Amérique du Nord, ceux-ci ayant été modifiés de différentes manières.[29]
Espèces exotiques [modifier]
L'introduction d'espèces invasives exotiques s'est produite à de nombreux endroits et pour de nombreuses raisons, dont le ballastage des navires de commerce. Un exemple très connu et bien étudié est l'introduction de la perche du Nil dans le lac Victoria. À partir des années 1960, la perche du Nil a progressivement exterminé les 500 espèces de cichlidés que l'on ne trouvait nulle part ailleurs que dans ce lac ; certaines espèces ne survivent que grâce à des programmes de reproduction en captivité, mais d'autres sont probablement éteintes.[30] Parmi les espèces de poissons invasives ayant causé des problèmes écologiques, on peut noter les carpes, les tête-de-serpent, les tilapias, la perche européenne, la truite fario, la truite arc-en-ciel ou la lamproie marine.

Le
requin baleine, le plus grand poisson au monde, est classé comme « vulnérable ».
Statut de préservation [modifier]
En 2006, la liste rouge de l'UICN comprenait 1 173 espèces de poissons menacées d'extinction.[31] Cette liste incluait des espèces comme la morue de l'Atlantique, le Cyprinodon diabolis, les cœlacanthes ou le grand requin blanc. Comme les poissons vivent sous l'eau, ils sont plus compliqués à étudier que les animaux terrestres ou les plantes, et on manque toujours d'informations sur les populations de poissons. Les poissons d'eau douce semblent particulièrement menacés, car ils vivent souvent dans des zones restreintes.
Mesures de protection [modifier]
Les echelles à poissons [modifier]
Les quotas de pêche [modifier]
Le poisson dans la culture humaine [modifier]
Dans les récits fondateurs [modifier]

Gravure préhistorique de l'époque néolithique, montrant une silhouette mi homme, mi-poisson et de nombreux poissons
Dans la mythologie égyptienne [modifier]
- Le tilapia, poisson qui a coutume de mettre ses petits dans sa bouche en cas de danger et de les laisser ressortir une fois la menace passée, est associé à la déesse Hathor, emblème de la résurrection.[32]
- Lorsque Seth découpa le corps d'Osiris en 14 morceaux et les dispersa dans le monde, seuls 13 furent retrouvés. Le 14e (le sexe) avait été dévoré par un poisson.
Dans la mythologie gréco-romaine [modifier]
- Triton est une divinité marine, fils de Poséidon et d'Amphitrite. Il est figuré avec une tête et un buste d'homme et une queue de poisson.
Dans la mythologie celtique [modifier]
Dans le judéo-christianisme [modifier]
- Selon la tradition juive, Dieu servira aux justes Léviathan, poisson monstrueux, comme nourriture à la fin des temps
- Jonas, selon la bible (Jonas, 2:1), a passé trois jours et trois nuits dans le ventre d'un poisson : « Yahvé fit qu'il y eut un grand poisson pour engloutir Jonas. Jonas demeura dans les entrailles du poisson trois jours et trois nuits ».
- Après sa Résurrection, Jésus, sur les bords du lac de Tibériade, prépare sur la braise un repas de poissons pour les disciples. Déjà lors des multiplications des pains, il avait fait distribuer du poisson aux foules affamées.
- Très tôt le poisson deviendra un symbole majeur du christianisme primitif
- Le poisson est devenu en chrétienté le repas du vendredi, mais aussi le met de pénitence et du temps de carême, ainsi que la nourriture ordinaire des religieux.[33].
Dans les légendes du Moyen-Age [modifier]
- Les sirènes était des êtres mi-femmes, mi-poissons.
La constellation des Poissons [modifier]
Dans les réalisations artistiques [modifier]
Le Cauchemar de Darwin (Darwin's Nightmare), film documentaire de Hubert Sauper
Dans la bande dessinée [modifier]
Quelques records [modifier]
Le paiche d'Amazonie, ou Arapaïma, est le plus gros poisson dulçaquicole : il peut atteindre 4,5 m et 200 kg. Le plus gros poisson connu est le requin baleine (16 m).
Selon les Proceedings of the Royal Society (janvier 2006), le plus petit poisson serait Paedocypris progenetica, un cyprinidé d'eau douce découvert par Maurice Kottelat et Tan Heok Hui, du Raffles Museum (Singapour). La femelle Paedocypris est mature à partir de 7,9 mm de long. Adultes, les femelles mesurent 10,3 mm et les mâles, 11,4 mm. Il détrône un gobie nain de l'océan Indien décrit en 1981, Trimmatom nanus, adulte à 8 mm. Un poisson du même genre, Paedocypris micromethes, légèrement plus grand (femelle adulte à partir de 8,8 mm), a été identifié à Sarawak. Ces poissons vivent dans les forêts marécageuses constituées d'arbres inondés poussant sur un sol de tourbe détrempée, mou et épais de plusieurs mètres. L'eau y est rouge sombre et très acide.
Selon une équipe de l’université de Washington, Photocorynus spiniceps est le plus petit vertébré connu : le mâle de cette espèce de baudroie de la famille des Cérates, découverte dans les abysses du large des Philippines, long de seulement 6,2 mm, vit en parasite sur le dos d’une femelle mesurant 46 mm. Celle-ci pourvoit aux besoins en nourriture d'un mâle ne se limitant pratiquement qu'à un appareil reproducteur (Pietsch et al., Ichtyological Research, 2005).
Si le mâle Photocorynus spiniceps est plus petit, notons qu'en moyenne Paedocypris progenetica détient le record du plus petit vertébré connu.
Dunkleosteus terrelli, un poisson marin à plaques cuirassées qui vivait il y a 400 Ma, mesurait jusqu'à 11 m de long et pouvait peser jusqu'à quatre tonnes. Après en avoir reconstitué la musculature, des scientifiques américains ont découvert en 2006 que les mâchoires de ce poisson étaient capables d'exercer une force de 5 500 kg/cm², soit à peu près deux fois celle de l'actuel requin blanc et autant que la mâchoire d'un tyrannosaure. De surcroît, Dunkleosteus terrelli était capable d'ouvrir et de refermer sa gueule en un cinquantième de seconde (Philip Anderson et Mark Westneat, 2006).

















